喜旱莲子草入侵对三峡库区重庆主城河岸带植物多样性的影响
时间:2022-12-10 09:55:03 来源:柠檬阅读网 本文已影响 人 
张丽苗,谭雪,董智,郑杰,袁中勋,李昌晓
(西南大学生命科学学院,三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,重庆 400715)
河岸生态系统作为陆地生态系统和水生生态系统之间的过渡带,对维持河道和流域的健康至关重要[1]。然而,由于干流和支流具有不同的河道河岸宽度和水系特征,因此对自然(河流侵蚀和沉积过程)和人为作用过程的响应存在差异[2],位于干流的河岸生态系统与支流的河岸生态系统往往表现出不同的植物群落特征。越来越多的研究表明,河岸生态系统是最容易受到生物多样性变化影响的区域之一[3]。尤其是在城市区域,城市河流及其河岸环境受到许多生物和非生物因素的影响,包括外来物种入侵、水位波动、富营养化、河堤建设、商用码头以及农业耕种等[4-5]。河流生态系统中的水位波动有助于外来物种的入侵和扩散,进而改变群落组成、结构和多样性[6-8]。外来物种的入侵已经成为一个主要的生态环境问题和生态学研究焦点[9-11],但学者们对于城市生态系统中的物种入侵研究却较少[12]。因此,城市河岸带作为引入物种进一步扩散到另一区域的主要来源,对其进行管理至关重要[13]。
喜旱莲子草(Alternanthera philoxeroides)又名空心莲子草、空心苋等,原产于南美洲的巴西、乌拉圭、阿根廷等国[14],于20世纪30年代末作为马饲料引入中国上海,后传播到华东、华中、华南和西南等广大地区。2003年名列国家环保总局公布的“中国第一批外来入侵物种名单”。该草为多年生宿根植物,水旱两栖均可生长。因具有极强的生境适应性、表型可塑性和无性繁殖适应性,导致其在入侵地能够迅速繁殖蔓延,常形成单一的优势群落[15-16],难以控制;
常使本地植物生长空间受限,危害生物多样性[17],破坏生态系统。三峡库区建立后,由于周期性的水位涨落,河岸带植物群落呈现出陆地构建-水淹退化-再构建的循环演替特征[18-19]。在此过程中,大量外来物种入侵,尤其是喜旱莲子草,占据着优势地位[20]。
近年来,已有许多学者对三峡库区外来物种入侵与景观基质组成结构的关系[19],城市河岸系统外来物种的入侵[12,21]以及外来物种入侵与环境因子间的关系[22-24]进行研究,但很少有人关注入侵物种对城市河流干流河岸带和支流河岸带物种多样性和物种分布特征的影响。因此,本研究旨在探讨喜旱莲子草入侵对城市河流干流和支流河岸带物种多样性的影响及其土壤决定因子。并提出以下科学问题:1)干流河岸带和支流河岸带的物种多样性是否存在差异?2)随着喜旱莲子草入侵强度的变化,干流河岸带和支流河岸带的物种多样性呈现怎样的变化趋势?3)影响干流河岸带和支流河岸带物种多样性和分布特征的土壤因子有何不同?为了回答这些问题,本研究对干流河岸带和支流河岸带的植被群落进行了大规模调查研究。
1.1 研究区概况
研究区位于三峡库区重庆主城(29°26′-29°37′N,106°22′-106°37′E)(图1),地处长江干流和嘉陵江汇流地带,地貌以丘陵和低山为主,海拔高度168~400 m,属亚热带季风性湿润气候,全年日照总数1000~1400 h,年平均气温18~20℃,年平均降水量1000~1350 mm[25-26]。该地区境内长江干流自西向东横贯全境,支流水系发达。由于三峡大坝的建设,库区呈现出冬蓄夏排的特殊水文节律,导致河岸植被生物多样性严重降低和外来物种入侵,如喜旱莲子草,小蓬草(Conyza canadensis)和鬼针草(Bidens pilosa)等[20]。
图1 研究区位置及样方分布Fig.1 Location of the study area and the investigated plots
1.2 样品采集和处理
2020年8月,在长江干流(寸滩干流、木洞干流、沙溪干流)和7条支流(双溪河、一品河、花溪河、五布河、双河、嘉陵江、御临河)河岸带进行植物群落调查。因三峡库区重庆主城段支流水系发达,分布广,面积大[2],故本研究对其增加采样强度。在前期踏查的基础上,选取47个典型代表性样点(干流12个、支流35个),样点区域为100 m×20 m,平行于河流。在每个样点内以50 m的间隔对3个2 m×2 m的取样样方进行全面调查,记录每个样方中的物种多度、盖度、高度和频度,以评估研究区域的群落结构和物种组成[27-28]。同时,测量每个样方的环境信息,以呈现地形变量。用五点取样法从样方0~20 cm深度处采集表土样品,各点混合均匀后用四分法取1 kg土壤带回实验室测量土壤理化性质。使用便携式GPS设备(UniStrong Odin系列,北京)在每个样带记录地理位置(纬度和经度)。
1.3 土壤理化特性的测定
采用烘干法测定土壤含水量(soil moisture,SM);
采用环刀法测定土壤容重(bulk density,BD);
采用氧化还原电位测定仪(HI98121,意大利)测定土壤温度(soil temperature,T)和氧化还原电位(redox potential,ORP);
采用水浸提液电位测定法测定pH;
采用重铬酸钾容量法-稀释热法测定有机质(soil organic matter,SOM)含量[29];
采用元素分析仪(elementar vario EL cube,德国)测定全氮(total nitrogen,TN)和全碳(total carbon,TC)含量;
采用电感耦合等离子体发射光谱法测定全磷(total phosphorus,TP)和全钾(total potassium,TK)含量;
采用全自动间断化学分析仪(CleverChem Anna,德国)测定速效磷(available phosphorus,AP)、速效钾(available potassium,AK)、铵态氮(ammonium nitrogen,NH4+-N)和硝态氮(nitrate nitrogen,NO3--N)含量[30]。
1.4 数据计算
重要值(importance value,IV)是度量植物生长及优势度的综合指标,重要值=(相对盖度+相对多度+相对频度)/3[19];
某种植物的总重要值为所有样地中该物种的相对重要值总和[31]。采用α-物种多样性指数度量样地群落的植物多样性水平,公式参考张金屯[32]的方法。

式中:S为样地内所有植物的物种数,Pi为物种i的重要值。
1.5 数据分析
使用SPSS 22.0软件对干流河岸带和支流河岸带喜旱莲子草的相对重要值、物种多样性及土壤因子差异进行独立样本t检验。根据喜旱莲子草盖度大小划分为4个入侵强度等级:无入侵(盖度0~0.1),轻度入侵(盖度0.1~0.3),中度入侵(盖度0.3~0.6),重度入侵(盖度0.6~1.0)[33]。对喜旱莲子草入侵强度与物种多样性进行回归分析,以软件提供的曲线方程建立模型,选择回归显著且拟合系数最高者进行讨论。
为了探讨土壤因子对物种多样性和物种分布的影响,首先使用Canoco 5(Microcomputer Power,美国)软件进行去趋势对应分析(detrended correspondence analysis,DCA),通过排序检验使用线性模型或单峰模型。如果DCA分析结果中排序轴第一轴长度小于3.0,使用基于线性模型的冗余分析(redundancy analysis,RDA);
如果大于4.0,使用基于单峰模型的典范对应分析(canonical correspondence analysis,CCA);
如果介于3.0~4.0,则二者皆可[34]。同时,在分析前通过方差膨胀因子(variance inflation factor,VIF)剔除共线性强的土壤因子。结果表明,优先选择RDA检验土壤因子对物种多样性的影响,选择CCA检验土壤因子对物种分布(选择总IV>1.0的物种)的影响。采用交互前向选择(forward selection)的方法选择最优环境变量,解释贡献率最大的变量表示对物种多样性和分布的影响最大[31]。
2.1 干流和支流河岸带植物物种组成与多样性
本研究共调查到77种植物,隶属于29科64属。其中,干流河岸带有20种,隶属于8科18属,禾本科、蓼科和菊科是干流河岸带的优势科,占70%;
支流河岸带有76种,隶属于29科64属,禾本科、菊科和唇形科是支流河岸带的优势科,占40.26%。
从优势种组成来看,在干流河岸带,牛鞭草(Hemarthria altissima)的 总 重 要 值 最 大(IV=14.45),其 次 是 狗 牙 根(Cynodon dactylon)(IV=8.80)和喜旱莲子草(IV=4.17)(表1);
在支流河岸带,喜旱莲子草的总重要值最大(IV=27.05),其次是狗牙根(IV=14.22)和牛鞭草(IV=11.34)(表2)。

表1 干流河岸带调查物种名录Table 1 Surveyed species list in the main waterway riparian zone

表2 支流河岸带调查物种名录Table 2 Surveyed species list in the tributary riparian zone
独立样本t检验表明,干流河岸带喜旱莲子草的平均相对重要值显著低于支流河岸带(P<0.05)。干流河岸带Patrick丰富度指数(P<0.05)和Shannon-Wiener多样性指数(P<0.05)显著低于支流河岸带,但干流河岸带Pielou均匀度指数显著高于支流河岸带(P<0.05),Simpson多样性指数(P=0.432)在干流和支流河岸带之间差异不显著(图2)。
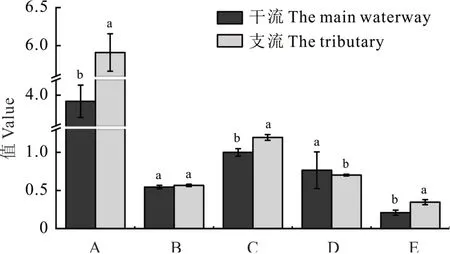
图2 干流和支流河岸带物种多样性及喜旱莲子草平均相对重要值比较Fig.2 Comparison of species diversity and average relative importance value of A.philoxeroides between the main waterway and tributary riparian zone
2.2 喜旱莲子草入侵对干流和支流河岸带物种多样性的影响
回归分析结果表明,喜旱莲子草的入侵强度(盖度)与Simpson多样性指数(干流:R2=0.3559,P<0.05;
支 流 :R2=0.3957,P<0.0001)、Shannon-Wiener多样性指数(干流:R2=0.2230,P<0.05;
支流:R2=0.2898,P<0.0001)以及Pielou均匀度指数(干流:R2=0.5837,P<0.05;
支流:R2=0.3603,P<0.0001)均有显著的二项式回归关系(图3)。随着入侵强度的增加,这3个多样性指数呈先上升后下降的单峰变化趋势,即小规模的喜旱莲子草入侵在一定程度上可以促进群落物种多样性和均匀性,但干流与支流的入侵盖度阈值耐性指数不同(如图3箭头所示:干流 的Simpson多 样 性指 数0.45,Shannon-Wiener多样性指数0.77,Pielou均匀度指数0.45;
支流的Simpson多 样 性 指 数0.33,Shannon-Wiener多 样 性指数0.18,Pielou均匀度指数0.38)。喜旱莲子草入侵对干流河岸带物种丰富度指数(R2=0.0240,P=0.816)无显著影响,但与支流河岸带物种丰富度指数(R2=0.1600,P<0.0001)有显著的负相关关系。
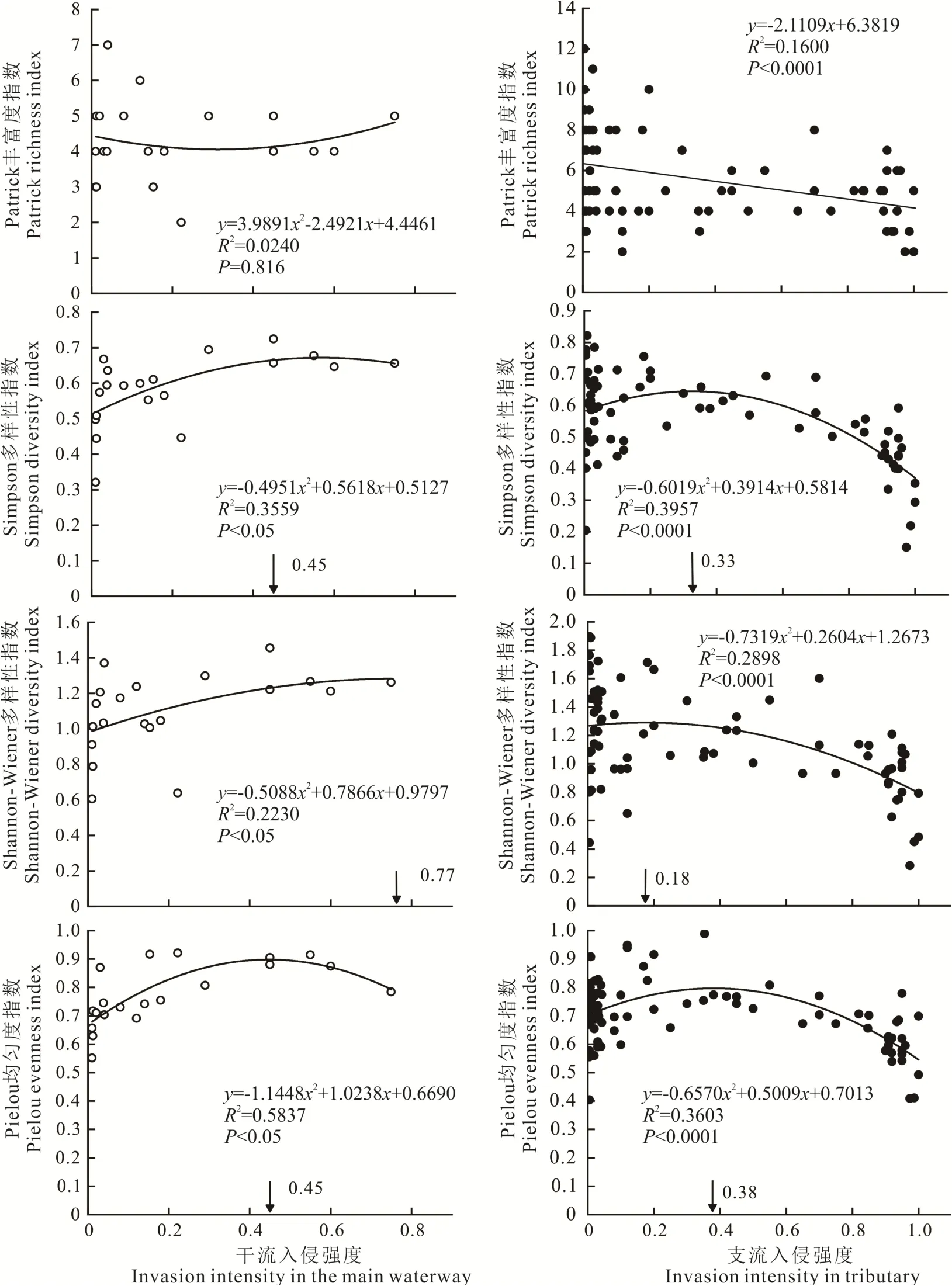
图3 喜旱莲子草入侵强度对干流和支流河岸带物种多样性的影响Fig.3 The effect of A.philoxeroides invasion on species diversity in the main waterway and tributary riparian zone
2.3 干流和支流河岸带土壤因子的差异
独立样本t检验表明,干流河岸带土壤pH和温度显著高于支流河岸带(P<0.05),而土壤全氮、全钾、有机质、铵态氮、速效磷、速效钾含量及氧化还原电位显著低于支流河岸带(P<0.05);
土壤全碳、全磷、容重、硝态氮及含水量在干流和支流河岸带之间差异不显著(P>0.05)(表3)。

表3 干流和支流河岸带土壤因子特征Table 3 Characteristics of soil factors in the main waterway and tributary riparian zone
2.4 土壤因子对干流和支流河岸带物种多样性的影响
RDA结果表明,在干流河岸带,物种多样性与环境因子关系的前两个轴的累积解释率为40.04%(图4a),在支流河岸带,前两个轴的累积解释率为46.21%(图4b)。前向选择表明,在干流河岸带,土壤有机质的解释率和贡献率(21.1%和51.7%)最大,硝态氮和全氮的解释率和贡献率(分别为6.3%、15.5%和3.7%、9.1%)也较大(表4)。而在支流河岸带,土壤含水量的解释率和贡献率(17.9%和41.7%)最大,全氮和速效磷的解释率和贡献率(分别为9.6%、22.4%和5.1%、11.8%)也较大(表4)。
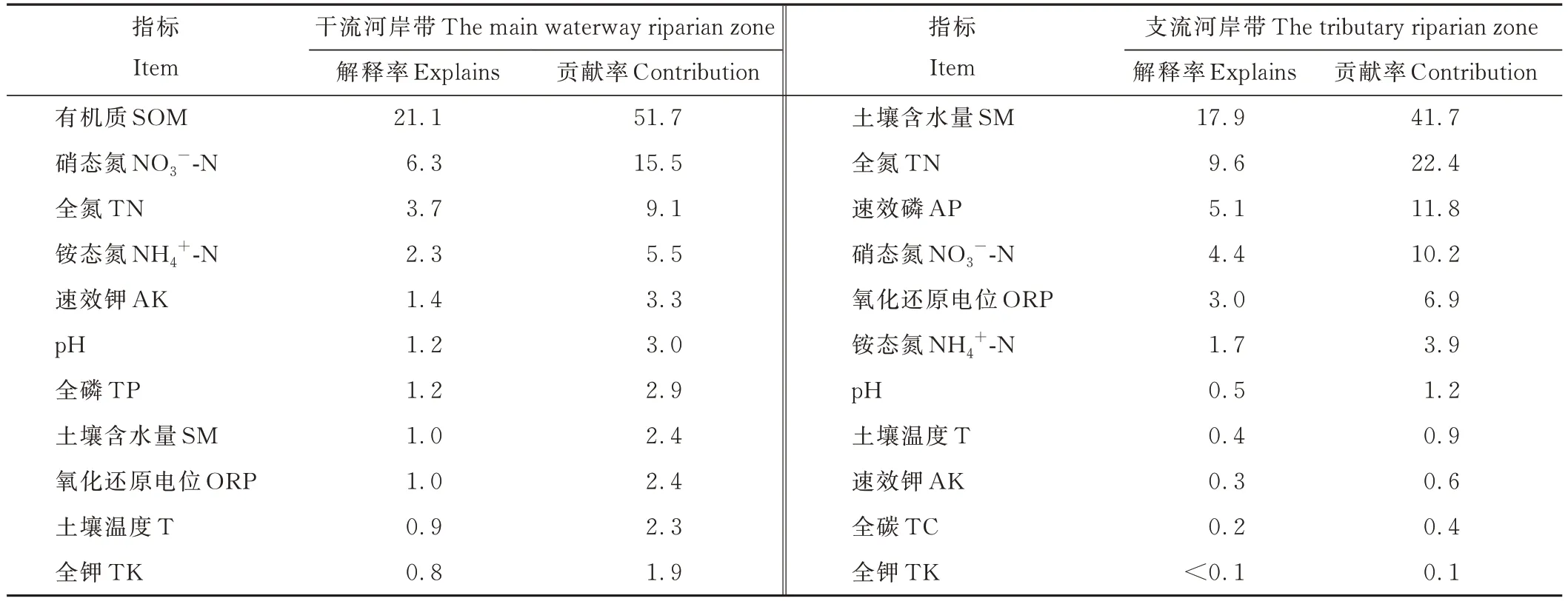
表4 物种多样性指数进行RDA分析的前向选择Table 4 Forward selection for the RDA on diversity index(%)
土壤因子对干、支流河岸带物种丰富度的影响存在差异。土壤温度、pH及氧化还原电位对支流河岸带物种丰富度和多样性指数有一定的促进作用,但是抑制干流河岸带物种丰富度和多样性指数。土壤全氮、速效钾及含水量对干、支流河岸带物种丰富度和多样性的影响正相反。在干流河岸带,土壤有机质和含水量与喜旱莲子草盖度正相关,土壤pH、温度和氧化还原电位与喜旱莲子草盖度负相关。在支流河岸带,土壤速效磷、铵态氮、全碳和含水量与喜旱莲子草盖度正相关,土壤硝态氮与喜旱莲子草盖度负相关。此外,土壤速效钾、全氮和全钾与干、支流河岸带喜旱莲子草盖度均正相关(图4)。
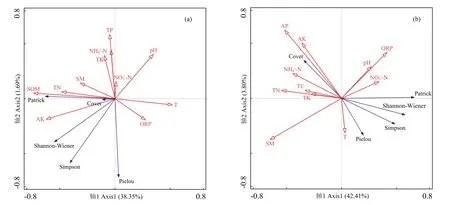
图4 干流(a)和支流(b)河岸带物种多样性与环境因子的RDA分析Fig.4 RDA analysis of species diversity and environmental factors in the main waterway(a)and tributary(b)riparian zone
2.5 土壤因子对干流和支流河岸带物种分布的影响
CCA结果表明,在干流河岸带,物种分布与环境因子关系的前两个轴的累积解释率为69.00%(图5a),在支流河岸带,前两个轴的累积解释率为73.73%(图5b)。前向选择表明,在干流河岸带,土壤速效钾的解释率和贡献率(12.2%和26.0%)最大,含水量和pH的解释率和贡献率(分别为8.7%、18.6%和5.7%、12.2%)也较大(表5)。喜旱莲子草(3)分布在第一象限,受土壤速效钾、有机质和全氮的影响较大(图5a)。而在支流河岸带,速效磷的解释率和贡献率(3.8%和17.7%)最大,土壤含水量和温度的解释率和贡献率(分别为2.8%、12.9%和2.8%、12.8%)也较大(表5)。喜旱莲子草(1)分布在第一象限,受土壤速效磷、速效钾和全氮的影响较大(图5b)。
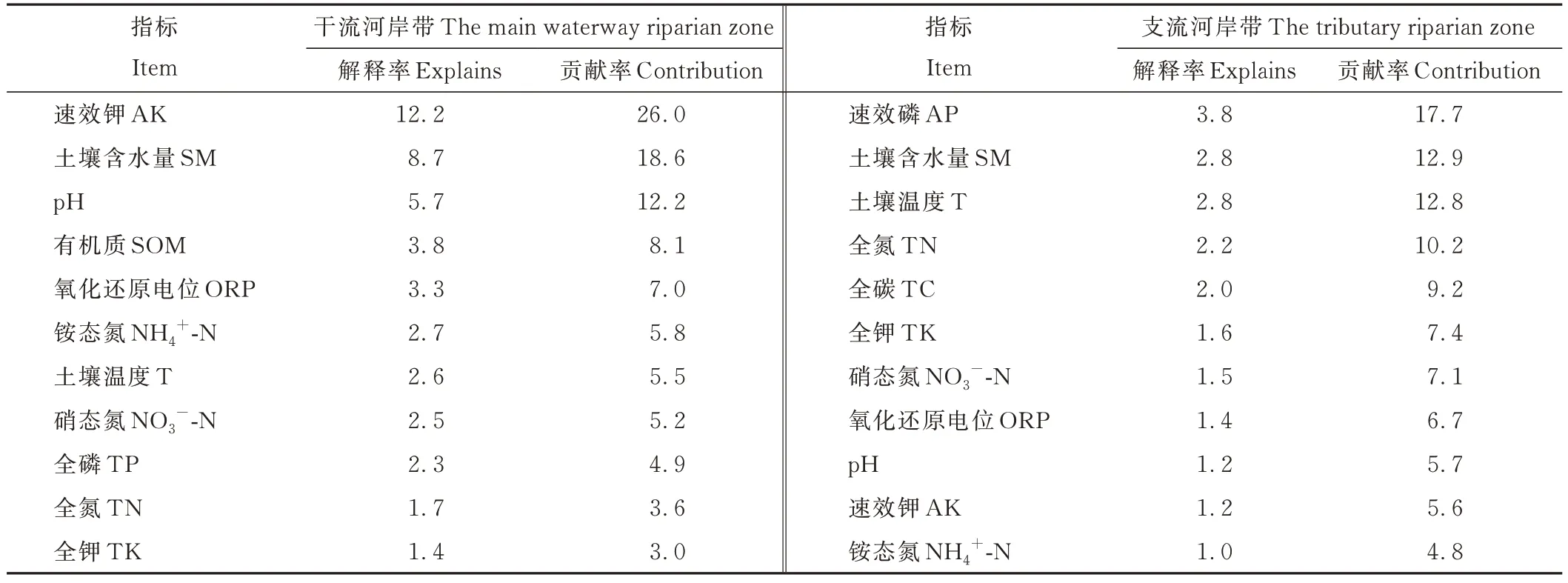
表5 物种分布进行CCA分析的前向选择Table 5 Forward selection for the CCA on species distribution(%)
随着全球气候变化和经济全球化的迅速发展,外来物种入侵对生态系统功能和全球生物多样性构成了越来越严重的威胁[35-36],已成为当代社会面临的最严峻的生态问题之一[37]。2021年10月在昆明召开的联合国《生物多样性公约》第15次缔约大会(15th Conference of the Parties,COP15)足以证明生物多样性的重要。城市河岸带作为提供高度生物多样性和重要生态系统服务的生境,易成为外来物种入侵的载体[38]。本研究表明三峡库区重庆主城河岸带物种多样性正受到喜旱莲子草入侵的破坏。
本研究发现干流河岸带Patrick丰富度指数和Shannon-Wiener多样性指数显著低于支流河岸带,但干流河岸带Pielou均匀度指数显著高于支流河岸带,Simpson多样性指数在干流和支流河岸带之间差异不显著。由于三峡库区干流和支流具有不同的河道河岸宽度和水系特征,因此在发生水文节律变化时,河流侵蚀和沉积过程不同[39-40]。并且,此次调查是在洪水过后的平水期进行,不稳定的水流循环对土壤进行剧烈冲刷,使河岸带土壤养分缺失。同时,洪水也阻碍了很多不耐水淹物种的再生甚至导致其消失。河岸带主要为洪水过后形成的河漫滩,通常是耐水淹的优势物种形成的优势群落。在较宽的干流河岸带,人为干扰多远离水体,而支流河岸带易获得因农田干扰、人类活动及边缘效应等传播的种子,使河岸带物种多样性增大[2,41]。研究表明硝态氮有利于植物生长,而高浓度的铵态氮可能会减少植物对于碱性离子的摄取从而降低其阳离子吸附能力,影响根系的正常生长,对植物产生毒害作用[31]。本研究发现硝态氮对支流河岸带物种丰富度和多样性指数有一定的促进作用,而对干流河岸带的影响正相反,铵态氮对干、支流河岸带的影响均是负面的。因此,干流河岸带的物种丰富度和多样性指数较低。但干流河岸带Pielou均匀度指数显著高于支流河岸带,可能是因为支流河岸带土壤有机质高而pH低,入侵物种通过其强大的竞争力,干扰了支流河岸带的异质性,并大幅度降低了物种均匀度,从而导致支流河岸带物种均匀度较低[42],这与Wu等[31]的研究结果一致。
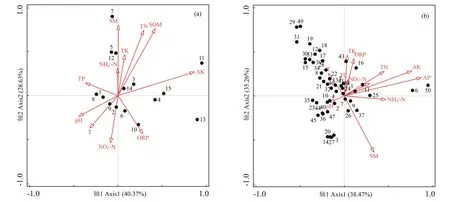
图5干流(a)和支流(b)河岸带物种分布的CCA分析Fig.5 CCA analysis of species distributions in the main waterway(a)and tributary(b)riparian zone
本研究量化了喜旱莲子草不同入侵强度与物种多样性之间的关系。生物入侵通常会降低全球范围内的生物多样性,但会提高区域范围内的物种多样性[43]。本研究发现小规模的喜旱莲子草入侵会促进本地生态系统物种多样性,这在先前有关外来物种入侵的研究中得到证实[44-45]。而大规模的喜旱莲子草会降低本地生态系统物种多样性,这与先前的研究结果一致[46]。可能是因为在入侵初期,繁殖压力高,资源竞争压力低[47],喜旱莲子草作为具备优越的表型可塑性和生理适应特性的外来物种[48],迅速建群并改善土壤性能和栖息地的适宜性,因此物种多样性和群落可入侵性都有一个上升的趋势。然而,随着喜旱莲子草的生长,其形成密集的根茎结构并释放出化感物质影响周围物种的生长[49],并通过资源竞争和物种相互作用,导致物种多样性降低[50-51]。同时,本研究发现支流河岸带中喜旱莲子草的平均相对重要值显著高于干流河岸带,且Simpson多样性指数、Shannon-Wiener多样性指数及Pielou均匀度指数的喜旱莲子草入侵盖度阈值均低于干流河岸带,表明支流河岸带更容易受到喜旱莲子草入侵的影响。可能是因为支流河岸带坡度较小,水流流速低[2],相对于干流河岸带更多耐水淹的牛鞭草和狗牙根来说,喜旱莲子草更容易在土壤有机质、全氮、速效磷、速效钾含量高而pH低的支流河岸带小生境中成为优势种。这一结果与Chen等[52]和王桔红等[53]的结论一致,即土壤有机质和全氮含量较高的河岸带,易受喜旱莲子草入侵,喜旱莲子草的盖度更高。Daehler[54]有关生境可入侵性的研究也表明,外来物种更容易入侵土壤养分较高的生境或群落。
河岸带通常在区域范围内具有高度异质性[28]。植物对洪水的冲刷和掩埋、缺氧和水淹的耐受性随河岸带环境的变化而变化[50-51],群落属性(物种丰富度、多样性、盖度)、植被覆盖类型以及植物功能分组也会对此做出相应的响应[55]。土壤有机质、氮、磷、钾等元素作为植物生长和发育必需的营养元素,对于维持物质循环平衡具有重要意义[52]。本研究通过对干流河岸带和支流河岸带物种分布特征与土壤因子关系的分析发现,土壤速效钾、速效磷、含水量、pH和温度是影响河岸带物种分布特征的主要因子。喜旱莲子草入侵通过和其他物种竞争养分吸收,改变进入土壤中的凋落物的数量和质量,或间接通过根系活动而改变入侵生态系统中的土壤营养循环[56]。有机质、速效磷、速效钾等养分含量及pH、温度也随之发生变化,从而影响植物的养分利用率[57]。土壤性质的变化反过来又会影响群落的物种组成[24]。关于河岸带物种分布特征与环境因子的关系,不同地区的研究结果不尽相同。张志永[58]的研究结果表明,影响三峡库区河岸植物群落特征的主要环境因子是高程和土壤有机质含量。桂东伟等[59]的研究结果表明,影响中昆仑山北坡策勒河植被的主要环境因子是海拔、pH、有机质和土壤含水量等。河岸带微生境复杂,土壤养分含量、植物种源特征等共同决定着局部尺度上的植物群落组成。因此,研究植物分布与环境因子的关系具有重要的理论价值和实践意义。
通过对三峡库区重庆主城河岸带喜旱莲子草入侵与植物群落物种多样性的关系研究发现,随着喜旱莲子草入侵强度逐渐增加,Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数呈现出先升高后降低的趋势。喜旱莲子草的入侵在干流河岸带主要与速效钾、有机质和全氮含量有关,而在支流河岸带主要与速效磷、速效钾和全氮含量有关,并且喜旱莲子草的入侵程度表现为支流大于干流。对于城市河岸带,探索喜旱莲子草入侵的土壤环境决定因素对于预测其未来在河岸带的分布和优先管理工作至关重要。同时,在对河岸带进行恢复治理时,不能忽视支流河岸带。
猜你喜欢 支流干流全氮 清代珠江沿江城市空间分布及特征史学集刊(2021年5期)2021-09-11三峡库区支流“水华”现状及防控对策安徽农业科学(2019年3期)2019-06-11广西主要支流柳江治理工程(鹿寨县导江乡段)护岸建筑物型式设计科学与财富(2018年13期)2018-06-13南京市某城市内河支流水环境问题浅析安徽农学通报(2017年24期)2018-01-12西藏主要农区土壤养分变化分析农业与技术(2017年12期)2017-07-11三峡库区消落带紫色土颗粒分形的空间分异特征江苏农业科学(2016年6期)2016-07-25武功山山地草甸土壤有机质、全氮及碱解氮分布格局及关系江苏农业科学(2015年11期)2016-01-27套种绿肥对土壤养分、团聚性及其有机碳和全氮分布的影响天津农业科学(2015年9期)2015-09-02三峡成库后典型支流航运条件及通航管理对策研究建筑科技与管理(2008年10期)2008-03-30







